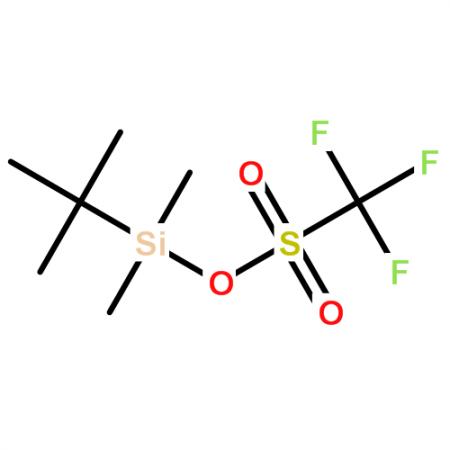




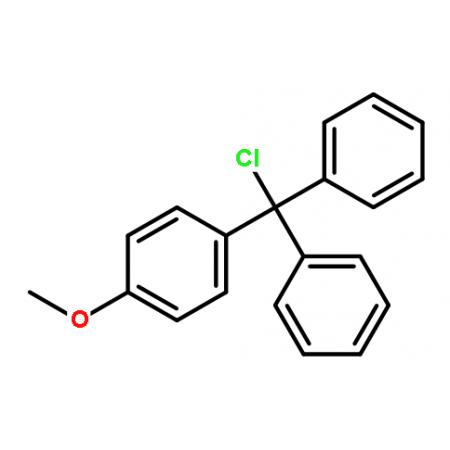










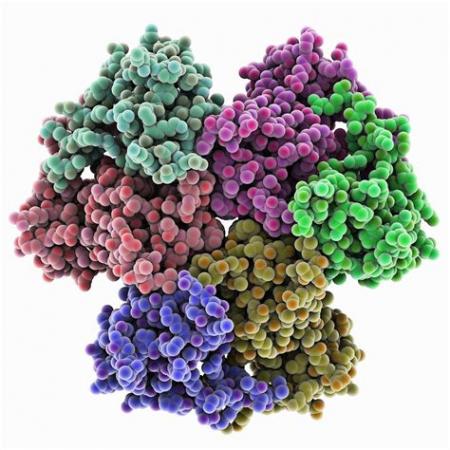


Muller等采用NMR结合局城性分子动力学计算研究了MMOR的结构[]表明,还原酶(sMMOR)有2个城,一个是含有FAD和NADH结合部位MMOR-FAD城,分子量为27. 6kDa,由250(99~348)个氨基酸组成;另一个是含有[2Fe-2S] 的铁氧化还原蛋白城(MMOR-Fd), 由98(1~98)个氨基酸组成,6个β带(strands), 3个a螺旋构成了2个β片, [2Fe-2S]中心与42、47、50、82半胱氨酸残基的硫原子配位,分子量为10. 9kDa,MMOR总分子量为38. 5kDa; MMOR-Fd从NADH向MMOH转移电子,Fd与MMOH结合面有一个极性中心,这个中心被一些疏 水基团包围。


Copyright© 2019-2029,先河原料科技(三明)有限公司版权所有
有任何购物问题请联系我们在线客服 | 电话:0598-7509639| 工作时间:周一至周六 8:00-18:00
【注】本站销售的产品【化学品】仅用于工业应用或者科学研究、化学分析、高精密电子清洗、科研机构实验、教学实验、农残检测、疾病控制(含消杀)、质量检测、医药中间体(ROHS)环保检测、水处理检测、日用化学品的研发与试剂检测、食品检测、石油化工涂料检测、工矿企业生产及检测等【非医疗目的】,不可用于人类或动物的临床诊断或治疗【非药用】【非食用】
![]() 闽ICP备15011996号-4
|
闽ICP备15011996号-4
|
![]() 闽公网安备35040002000218号
|
闽公网安备35040002000218号
|
![]() 营业执照(三证合一)
|
营业执照(三证合一)
|
![]() 危险品经营许可证
|
危险品经营许可证
|
![]() 易制毒经营备案
易制毒经营备案
